پیوند درخت عبارت است از اتصال و اتحاد دو اندام و بافت گیاهی بهطوریکه دو قسمت در محل اتصالیترمیم شده و بهصورت یک واحد مستقل درمیآیند. قسمتی که پیوند در روی آن انجام میگیرد و بخش ریشه گیاه جدید را تشکیل میدهد، پایه خوانده میشود و قسمتی که روی پایه پیوند میزنیم و نهایتاً اندام هوایی و شاخسار را تشکیل میدهد پیوندک نامیده میشود .
مزایای پیوند درختان میوه
با استفاده از پیوند، میتوان گیاهی را که دارای ریشه ضعیفی است روی پایهای که دارای ریشه قوی است قرارداد .
با پیوند زدن میتوان درختانی همانند بادام، سیب و گرد را که افزایش آنها با سایر روشها (مانند خوابانیدن) مشکل است؛ بهآسانی و به سرعت تکثیر نمود .
با پیوند میتوان ارقام جدید را با استفاده از پایههای قدیمی جایگزین ارقام قدیمی کرد. این کار را سرشاخه کاری میگویند .
با استفاده از پیوند میتوان نسبت به تعویض رقم باغ قدیمی اقدام کرد .
از دیگر مزایای پیوند جلو انداختن گلدهی و زود باردهی نسبت به سایر روشها میباشد .
عوامل موثر در پیوند موفق
زمان انجام پیوند
روش انجام پیوند
سازگاری بین پایه و پیوندک
شرایط محیطی
زمان پیوند
در پیوندهای شاخه بهترین زمان برای انجام پیوند اواخر زمستان – اوایل بهار قبل از بیدارشدن درخت از خواب زمستانی است. اما برای پیوند جوانه برحسب منطقه از بهار تا تابستان یعنی زمانی که پوست پایه بهراحتی جدا میشود انجام میگردد .
روشهای انجام پیوند
پیوند اسکنهای: پیوندی است که در اواخر اسفندماه تا اوایل فروردینماه در درختان میوه مورداستفاده قرار میگیرد .
پیوند نیمانیم یا پیوند انگلیسی: این نوع پیوند زمانی به کار میرود که پایه و پیوندک دارای قطر مساوی باشد و به دو صورت نیمانیم ساده و یا نیمانیم زبانهدار مورداستفاده قرار میگیرد .
پیوند جوانهای: این نوع پیوند همیشه در نهالهای جوانی که سن آنها بین ۱ تا ۲ سال است انجام میشود . این نوع پیوند را در سه موقع از سال میتوان انجام داد:
الف) فصل بهار (اواخر بهار تا اوایل تابستان)
ب) در اواسط تا اواخر تابستان نیز میتوان این پیوند را انجام داد
ج) در بهار قبل از بیدارشدن گیاه نیز میتوان این پیوند را انجام داد ودراین زمان همپایه و هم پیوندک بهخوبی باهم پوست میدهند .
پیوند سپری شکمی: این نوع از پیوند به دلیل سرعت عمل، سهولت جوش خوردن بافتها و ایجاد حداقل زخم در پایه بیشترین استفاده را دارد .
پیوند وصلهای: این روش بیشتر برای درخت گردو به کار میرود و زمانی انجام میشود که درخت بهراحتی پوست بدهد .
علائم ناسازگاری (نگرفتن) پیوند
اختلاف در رشد قطری پایه و پیوندک
تغییر رنگ زودهنگام برگها در پاییز بهاحتمال زیاد دلیل ناسازگاری و عدم گرفتن پیوند است .
ریزش غیرطبیعی برگ (ریزش زودهنگام برگ)
رشد سریع جوانههای گل و شکوفه کردن نابهنگام در پاییز میتواند از علائم ناسازگاری در پیوند باشد .
جداشدن پیوندک از پایه در مراحل اولیه رشد نهال
زرد شدن برگهای نهال در خزانه از دیگر علائم ناسازگاری است
بافتهای پیوندی در حکم یک فیلتر برای عبور مواد غذایی و مواد جذبشده از ریشه (املاح معدنی) است اما بعضی مواقع به دلیل ناسازگاری و نگرفتن پیوند از انتقال املاح معدنی جلوگیری شده و علائم کمبود عناصر غذایی ظاهر میگردد .
تشکیل برآمدگی در محل پیوند که به دلیل اختلاف در رشد پایه و پیوندک است .
پاکوتاهی و ضعف کلی درخت که باعث کوتاه شدن عمر درخت میگردد
عارضه خط سیاه که در پیوند گردوی ایرانی روی گردوی سیاه حتی پس از ۲۰ سال قابلمشاهده است .
عوامل محیطی مؤثر در جوش خوردن پیوند
دما: بعد از انجام عمل پیوند گرما و یا سرمای شدید باعث عدم موفقیت در پیوند میشود.
رطوبت: بعد از انجام پیوند به علت ازدسترفتن رطوبت، محل پیوند خشکشده و پیوند نمیگیرد؛ بنابراین برای جلوگیری ازدسترفتن رطوبت در محل پیوند باید از چسب پیوند استفاده نمود .
منابع
خوشخوی، م؛ روحانی، ا و تفضلی، ع . ۱۳۸۱. اصول باغبانی. انتشارات دانشگاه شیراز
رسولزادگان، یوسف . ۱۳۷۹. میوه کاری در مناطق معتدله. انتشارات دانشگاه اصفهان
در این مقاله به تغذیه درختان میوه با معرفی عناصر غذایی مورد نیاز گیاه و نحوه جذب انها خواهیم پرداخت در ادامه همراه ما باشید
عناصر غذایی موردنیاز گیاهان
تغذیه بهینه گیاه شرط اصلی بهبود کمی و کیفی محصولات است. در تغذیه گیاه و درخت نهتنها باید هر عنصر بهاندازه کافی در دسترس آن قرار گیرد، بلکه ایجاد تعادل و رعایت تناسب میان همه عناصر غذایی از اهمیت زیادی برخوردار است. عناصر غذایی موردنیاز گیاهان به سه دسته کلی تقسیم میشوند.
عناصر غذایی مضر
عناصری هستند که برای رشد و نمو گیاهان زیانآور هستند. حتی در برخی موارد غلظتهایی کم این عناصر میتواند موجب کاهش قابلتوجهی در عملکرد و رشد گیاه گردد. ازجمله این عناصر میتوان به سرب، کادمیوم، جیوه و نیکل اشاره کرد
عناصر غذایی مفید
عناصری هستند که در صورت وجود در محیط سبب بهبود رشد گیاه و یا گیاهان خاصی میشوند. برای مثال سدیم برای چغندرقند، سیلیس برای برنج و جو و یا کبالت و مولیبدن برای تثبیت بیولوژیکی نیتروژن مفید میباشند.
عناصر غذایی لازم یا ضروری
سه معیار برای ضروری بودن وجود دارد که عبارتاند از:
گیاه بدون آن عنصر قادر به تکمیل چرخه حیات خود نباشد.
وظیفه آن عنصر توسط عنصر دیگری قابل انجام و جایگزینی نباشد.
عصر مستقیماً در متابولیسم و تغذیه گیاه نقش داشته باشد.
عناصر لازم یا ضروری عبارتاند از:
عناصر پرمصرف
کربن اکسیژن و هیدروژن: ٦٠ تا ٩٠ درصد ماده خشک گیاهی را تشکیل میدهند و جز در موارد کمبود
آب کمبود آنها دیده نمیشود؛ و عمدتاً از طریق آبوهوا تأمین میشوند.
عناصر کودی شامل نیتروژن فسفر و پتاسیم
عناصر آهکی شامل کلسیم و منیزیم
گوگرد
عناصر غذایی کممصرف
لازم به ذکر است همانطور یکه گیاهان بدون عناصر غذایی پرمصرف قادر به ادامه حیات نیستند بدون عناصر کممصرف نیز قادر به ادامه حیات نخواهند بود. تفاوت عمدهای که با عناصر پرمصرف دارند به مقدار بسیار کمتر موردنیاز میباشند. میزان مصرف آنها برحسب قسمت در میلیون است درحالیکه میزان مصرف عناصر غذای پرمصرف بر اساس درصد است. این عناصر عبارتاند از آهن، روی، مس، بر، منگنز، مولیبدن و کلر میباشند.
ارزیابی وضعیت تغذیه درختان میوه
راههای مختلفی برای تشخیص کمبودها تعیین میزان عناصر غذایی قابلاستفاده در گیاهان وجود دارد از متداولترین این روشها میتوان به ١- آزمون خاک ٢ – آنالیزهای بافت برگ درختان ميوه ٣- تشخیص ظاهری عوارض تغذیهای اشاره کرد.
آزمون خاک
شامل سه مرحله نمونهبرداری صحیح، تجزیه دقیق عناصر و تفسیر صحیح نتایج تجزیه خاک است. آزمون خاک میتواند در گیاهان زراعی سبزی و صیفیجات یکساله کمک زیادی به تأمین نیازهای غذایی گیاهان نماید. بین آزمون خاک و تجزیه برگ در باغ ارتباط کمی وجود دارد. یعنی آزمون خاک نمیتواند راهنمای خوبی برای تعیین وضعیت تغذیهای درختان میوه باشد. آزمون خاک قبل از احداث باغ بهترین وسیله برای مشخص کردن بافت خاک، مقدار آهک خاک و غلظت عناصر کلسیم، منیزیم، پتاسیم و فسفر خاک است. در باغهای احداثشده آزمون خاک مکمل نتایج تجزیه برگی است و جهت مشخص نمودن ترکیب کودی باید مورداستفاده قرار گیرد. برای نمونهبرداری صحیح خاک نقاط دارای شرایط و نوع خاک یکسان مرزبندی شده و خاکهای متفاوت ازنظر بافت خاک، وضعیت زه کشی خاک، مقدار آهک خاک، عمق لایه سخت کفه زیرین بهصورت جداگانه نمونهبرداری میشود.
تجزیه برگ نشاندهنده مقدار عناصری است که توسط درخت جذبشده و به قسمتهای هوایی درخت منتقلشدهاند. تجزیه برگ تصویر نسبتاً کاملی از وضعیت تغذیهای درختان میوه ترسیم میکند. نتایج تجزیه برگ تا حدودی وضعیت عناصر غذایی درختان میوه را ترسیم میکند. تجزیه برگ برای باغهای دایر مفید است ولی برای احداث باغ میوه ابتدا باید خاک را تجزیه کرد.
در صورت کمبود ازت از برگهای پیر به برگهای جوان منتقلشده و علائم کمبود اکثر در برگهای پیر مشاهده میشود توقف رشد و زردی برگهای پایین درخت از علائم کمبود ازت است.
نیتروژن
توقف رشد برگها، کوچک شدن برگها و ارغوانی شدن برگهای پایینی
فسفر
رنگپریدگی سوختگی حاشیه برگها و زرد شدن حاشیه برگهای پایینی در درخت انگور سبب خشک شدن نوک خوشههای انگور میگردد.
زرد شدن پهنکبرگهای جوان و سبز باقی ماندن رگبرگهای آن، برگهای تازه روئیده زردی بیشتری نشان میدهند و درنهایت لکههای نارنجی بر روی برگهای انتهای توسعه مییابند.
بارزترین مشخصه کمبود روی ریز شدن برگها و جارویی شدن برگهای جوان در سرشاخه -مای درخت است وجود نقاط زرد کرمی درزمینهٔ سبز تیره برگهای میانی کمبود روی سبب غیر همزمان رسیدن میوهای انگور میشود.
روی
کمبود منگنز شبیه سایر ریزمغذیها در خاکهای آهکی اتفاق میافتد. برگها زرد کمرنگ میشوند و این زردی از حواشی برگ شروعشده و به سمت رگ برگ میانی توسعه می باد.
منگنز
کمبود بر در درختان میوه سبب کاهش رشد و نمو پرچمها، کاهش مدت گردهافشانی، سیاه شدن وسط میوه سیب و بدشکلی میوه میشود.
بُر
کمبود مس در درختان میوه سبب ایجاد شاخههای پر رشد با برگهای درشت به رنگ سبز تیره و با لکههای زردرنگ میشود. در صورت تشدید کمبود تمام شاخههای جوان خشکیده میشوند.
مس
خشکی برگها مخصوصاً حواشی آنها از علائم عمومی کمبود کلر در درختان میوه است. در مقایسه با کمبود کلر سمت آن گستردگی جهانی دارد.
کلر
رابطه علائم کمبود عناصر غذایی با تحرک آنها در درخت
محل ظهور علائم کمبود عناصر غذایی در گیاهان به میزان انتقال عناصر غذایی از برگهای پیر به قسمتهای جوان گیاه بستگی دارد. عناصر غذایی مانند نیتروژن، فسفر و پتاسیم بهراحتی از برگهای مسن به قسمتهای جوان گیاه جابجا میشوند. ازاینرو علائم کمبود این عناصر ابتدا در برگهای پیرتر گیاه دیده میشود. در مقابل، عناصر غذایی مانند کلسیم و منگنز تحت هیچ شرایطی از برگهای پیر به قسمتهای جوان منتقل نمیشوند. درنتیجه علائم کمبود این عناصر در قسمتهای جوان گیاه دیده میشوند.
میزان تحرک عناصر غذایی در داخل گیاه (عناصر کم تحرک و عناصر پر تحرک)
میزان تحرک عناصر در گیاهان
عنصر
متحرک
ازت
متحرک
فسفر
متحرک
پتاسیم
غیر متحرک
کلسیم
نسبتاً غیر متحرک
منیزیم
غیر متحرک
گوگرد
نسبتاً غیر متحرک
آهن
تحرک کم
روی
در شرایط کمبود نسبتاً غیر متحرک و در شرایط کفایت متحرک
مس
غیر متحرک
منگنز
نسبتاً غیر متحرک
بُر
تحرک متوسط
مولیبدن
متحرک
کلر
برای اطلاعات بیشتر در مورد عناصر مورد نیاز گیاه لطفا مقالات زیر را مطالعه فرمایید
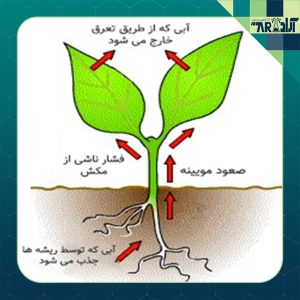
حرکت آب در گیاه : جهت رساندن آب و املاح جذب شده توسط ریشه به اندامهای هوایی و برگها، وجود یک سیستم انتقال ضروری است. این سیستم انتقال در گیاهان، شامل اجزای مختلفی میباشد. در این مقاله، ساختار سیستم انتقال آب در گیاه ، حرکت آب در گیاه و نیروی محرک در طی مسیر انتقال آب مورد بررسی قرار میگیرد.
ساختار سیستم هادی
آوندهای چوبی شامل چهار نوع سلول تراکئید (Tracheids)̨ عناصر وسل (Vessel)̨ فیبر (Fibers) و پارانشیم آوند چوب میباشند که از بین ساول ها تراکئید ها و وسل ها دارای آرایش عمودی بوده و در کار انتقال آب دخالت دارند. در آوند چوبی، مخصوصاً در درختان، فقط سلولهای پارانشیمی زنده هستند. البته سلولهای فیبر و پارانشیم در آوند آبکش نیز دیده میشوند.
سیستم آوند چوبی دارای انشعابها و شاخههای متعددی است که انتقال ایمن آب به شاخوبرگ گیاه را تضمین میکند، بدین ترتیب که اگر بخشی از آوندها صدمه ببیند، بقیه آنها کار انتقال آب را انجام میدهند.
وسل ها و ترکئید های سلولهای طویل و مردهای هستند که بعد از رشد و تمایز، بخشهای حیاتی آنها از بین رفته و پروتوپلاست آنها توسط سلولهای دیگر جذب میشود که ازنقطهنظر جریان آب مهم میباشد.
یکی از این تغییرات، تشکیل دیواره ثانویه است که عمدتاً از سلولز، لیگنین و همی سلولز تشکیل شده و بخش بزرگی از دیواره اولیه را میپوشاند.
تشکیل دیواره ثانویه، باعث استحکام وسل ها و تراکئید ها شده و از درهم ریختن آنها که معمولاً تحت مکشهای زیاد در آوند چوب به وجود میآید، جلوگیری میکند.
دیواره ثانویه لیگنینی شده مانند دیواره اولیه نسبت به آب نفوذناپذیر است، اما در آن منافذی به وجود میآید که از طریق آن منافذ دو سلول تراکئید یا وسل مجاور هم تنها توسط دیواره اولیه از هم جدا شدهاند.
نیروی محرکه در حرکت آب
دررابطهبا علت صعود شیره خام در آوند چوبی، چند تئوری مطرح است که شامل تئوری مویینگی، تئوری فشار ریشه و تئوری کوهیژن میباشد.
لازم به ذکر است که گرچه با قراردادن قطر منافذ دیوارهها ارتفاع صعود میتواند از نظر تئوری به سه کیلومتر نیز برسد، ولی ازآنجاییکه این منافذ بسیار ریز و میزان آب عبور کرده از آنها بسیار پایین است و از طرفی صعود آب به طریق مویینگی به چنین ارتفاعی تنها وقتی امکانپذیر است که آب خالص بوده و فاقد املاح باشد، درحالیکه آب موجود در خاک و آوند چوب خالص نیست، بنابراین، تئوری مویینگی پاسخگوی انتقال آب از آوند چوب نخواهد بود.
همچنین، ازآنجاییکه فشار ریشه تنها در شرایط خاصی که جذب اسمزی برقرار است، صادق بوده و برای ارسال آب برای درختان کافی نمیباشد، تئوری فشار ریشه نیز جهت انتقال آب از آوند چوب مناسب و کافی نیست.
بنابراین، امروزه نظریه پذیرفتنی، نظریه کوهیژن است.
ویژگیهای تئوری کوهیژن دررابطهبا مکانیسم انتقال آب در آوند چوب عبارتاند از:
دارای نیروی کوهیژن و استحکام کششی بالایی بوده و وقتی در لولهای باریک، مثل عناصر آوند چوب، قرار میگیرد میتواند کشش ۳ و حتی ۳۰ مگاپاسکال را قبل از پاره شدن ستون آب تحمل کند.
آب در گیاه، یک سیستم پیوسته را تشکیل داده و از دیوارههای سلولی اشباع از آب سلولهای برگ تا آوند چوب و از آنجا تا سطوح جذبکننده آب در ریشهها امتداد دارد. در واقع، یک طرف ستون آب در فضاهای بین سلولی دیواره سلولهای تبخیرکننده و طرف دیگر آن در فضای بین سلولی دیواره سلولهای جذبکننده آب میباشد.
تبخیر آب از سلولهای برگ، پتانسیل آب دیوارههای سلولی را کاهش داده و باعث میشود تا آب از آوند چوب به سطوح تبخیرکننده به حرکت درآید. این امر باعث کاهش فشار شیره آوند چوب شده و درنتیجه در سیستم هیدرولیک پیوسته گیاه، مکش ایجاد میشود.
کاهش پتانسیل آب در سطوح تبخیری به ریشهها منتقلشده و در آنجا سبب ورود آب از خاک به ریشه میشود؛ بنابراین، در گیاهان در حال تعرق جذب آب توسط سرعت تعرق کنترل میشود.
مقاومت در برابر حرکت آب
حرکت آب در طی مسیر جریان رابطه معکوسی با میزان مقاومت موجود در مسیر دارد. مقاومت میتواند در نقاط مختلف مسیر مانند ریشه، ساقه و برگ وجود داشته باشد، گرچه میزان آن در ریشه و برگ به مقدار قابلتوجهی بیشتر از ساقه است.
کارایی سیستم هادی
کارایی سیستم هادی باتوجهبه گونه گیاهی و شرایط رشد متغیر بوده و به طور معمول در موها بیشتر از گونههای گیاهی دیگر است. این مسئله به دلیل وجود آوندهایی با قطر بیشتر در موها است.
کنترل جریان
ازآنجاییکه بیش از ٥٩% آب جذبشده در اکثر گیاهان بهوسیله جریان تعرق تلفشده و تنها مقدار اندکی از آب جذبشده در گسترش سلول و فرایندهای بیوشیمیایی آن مصرف میرسد̨ بهخاطر کمی̨ جریان آب در گیاه توسط تلفات آب در تعرق تنظیم میگردد.
تبخیر آب از دیوارههای سلولی باعث ایجاد نیروی ماتریک یا فشار منفی یا جذبی در آنها شده و درنتیجه آب از آوند چوب بهسوی دیوارههای سلولی جاری میشود.
این امر، باعث ایجاد کشش یا فشار منفی در شیره آوند چوب شده̨ که به ریشه منتقل و درنتیجه جریان آب به داخل ریشه را ایجاد میکند.
بنابراین، افزایش تعرق، باعث افزایش جذب آب و کاهش تعرق، باعث کاهش جذب آب میشود.
در مقابل، گر جذب براثر عواملی مثل سرما، خشکشدن خاک و تهویه ناکامی کاهش یابد، پتانسیل آب ریشه کاهش مییابد.
کاهش پتانسیل آب ریشه، سبب افزایش مقاومت ریشه یا کاهش نیروی محرک شده و درنتیجه، تعرق کاهش مییابد. اگر گیاه بتواند پتانسیل فشاری منفیتری در برگها ایجاد کند، مجدداً تعرق بهسرعت قبلی خود بازمیگردد.
بههرحال درصورتیکه کاهش پتانسیل فشاری برگ بهقدری باشد که روزنهها بسته شوند̨ کاهش تعرق ایجادشده قابل بازسازی نیست.
بهطورکلی، جریان آب در گیاه بهوسیله تعرق کنترلشده و درواقع روزنهها تنظیمکنندگان اصلی جریان آب در گیاه هستند.
افزایش مقاومت در برابر حرکت آب، بهویژه در خاک و ریشه، تعرق را به طور غیرمستقیم و از طریق کاهش آماس برگ و بستهشدن جزئی تا کامل روزنهها کاهش میدهد.
بهعبارتدیگر، در شرایط مطلوب بودن رطوبت خاک، جذب تابع تعرق و در شرایط محدودیت رطوبت خاک، تعرق تابع جذب است که در نتیجه آن حرکت آب در گیاه بهبود می یابد.
در این مقاله قصد داریم به معرفی انواع کود کلات آهن موجود در بازار بپردازیم و به صورت مختصر به بررسی معایب و مزایای هر کدام بپردازیم
کمبود آهن در گیاهان
کمبود آهن یک عامل محدودکننده رشد گیاهان است. اگرچه این عنصر به میزان زیادی در خاک وجود دارد، اما قابلیت جذب آن توسط گیاهان معمولاً پایین است و بنابراین کمبود آهن یکی از مشکلات رایج در تغذیه گیاهی است.
قابلیت جذب آهن برای گیاهان
اگرچه مقدار زیادی از آهن موجود در پوسته زمین به شکل +٣Fe است، اما شکل +٢Fe ازنظر فیزیولوژیکی دارای اهمیت بیشتری برای گیاه است. این شکل از آهن معمولاً محلول بوده اما بهسرعت اکسیدشده و تبدیل به +٣Fe میشود. +٣Fe در pH خنثی و pHهای بالا نامحلول بوده و باعث میشود تا آهن در خاکهای قلیایی و خاکهای آهکی برای گیاه غیرقابلجذب باشد. علاوه بر این، در این نوع خاکها آهن بهراحتی با فسفاتها، کربناتها، کلسیم، منیزیم و یونهای هیدروکسید ترکیب میشود و از دسترس گیاه خارج میگردد.
معرفی کود های کلات آهن
جذب آهن توسط گیاه
گیاهان آهن را بهصورت های اکسیدشده آن جذب میکنند که شامل آهن فروس یا سهظرفیتی (+٣Fe) و آهن فریک یا همان دو ظرفیتی (+٢Fe) است. جذب آهن توسط گیاهان به روشهای مختلفی صورت میپذیرد که یکی از این روشها، کلاته کردن آهن است. در این روش، گیاه ترکیباتی به نام سیدروفور را از ریشه خود در خاک رها میکند که با آهن پیوند داده و حلالیت آن را افزایش میدهند. در این مکانیسم ممکن است باکتریها نیز دخیل شوند.
مکانیسم دیگری که گیاهان به کمک آن میزان جذب آهن خود را افزایش میدهند، آزادکردن پروتون (+H) در خاک و درنتیجه کاهش pH در محیط اطراف ریشه است که درنهایت منجر به افزایش میزان حلالیت آهن در خاک میشود. در اینجاست که انتخاب نوع کود نیتروژنه اهمیت بالایی پیدا میکند. شکل آمونیومی نیتروژن باعث افزایش آزاد شدن پروتون از ریشه شده و بنابراین باعث کاهش pH در خاک و افزایش جذب آهن میگردد. در مقابل، شکل نیتراتی نیتروژن موجب افزایش آزادسازی یونهای هیدروکسید شده و با افزایش pH خاک منجر به ایجاد تداخل در جذب آهن میگردد.
لازم به ذکر است که ریشههای جوان و تارهای کشنده (ریشههای مویین) توانایی بیشتری در جذب آهندارند و بنابراین داشتن سیستم ریشهای فعال و سالم از اهمیت بسزایی در تغذیه گیاهی برخوردار است. به طور خلاصه میتوان اینگونه بیان کرد هر عاملی که به رشد، توسعه و سلامت ریشه گیاه صدمه بزند در جذب آهن نیز اختلال ایجاد میکند.
معرفی کود های کلات اهن
مقابله با کمبود آهن
هنگامیکه علائم کمبود آهن در گیاه مشاهده میشود، میتوان با اسپری کردن آهنبر روی برگهای گیاه نسبت به برطرفکردن مشکل در کوتاهمدت اقدام کرد اما باید به یادداشت که در مورد گیاهان نیز همیشه پیشگیری بهتر از درمان است؛ بنابراین کشاورز باید دلیل بروز این کمبود را تشخیص داده و آن را برطرف کند تا این مشکل دیگر در آینده پیش نیاید.
در اغلب موارد، بروز علائم کمبود آهن به معنای ناکافی بودن مقدار آهن در خاک نیست بلکه ممکن است به دلیل شرایطی باشد که دسترسی گیاه به آهن موجود در خاک را محدود میکند. میزان کربنات در خاک، شوری خاک، رطوبت خاک، سرد بودن خاک و میزان سایر عناصر در خاک (بهعنوانمثال فسفر، کلسیم و عناصر کممصرف رقیب) ازجمله این شرایط هستند. بررسی این عوامل و برطرفکردن آنها تا حد ممکن، میتواند از هدررفت زمان و هزینه برای مصرف کودهای آهن که شاید نیازی به آنها نباشد جلوگیری کند.
کودهای آهندار
کودهای آهندار به دو صورت سولفات آهن و یا شکلهای کلاته شده ارائه میشوند.
سولفات آهن (٤FeSO)
این کود معمولاً دارای ٢٠% آهن است. سولفات آهن کودی ارزانقیمت است که هنگامیکه در خاکهای نامساعد مصرف شود بهسرعت غیرفعال میگردد. این وضعیت در pHهای بالای ۷ بحرانیتر است زیرا آهن موجود در کود بهسرعت تبدیل به +٣Fe شده و برای گیاه غیرقابلجذب میشود.
کلاتهای آهن
کلاتها ترکیباتی هستند که یونهای فلزی (در اینجا آهن) را تثبیت کرده و از اکسید شدن آنها جلوگیری میکنند.
کلاتهای آهن از سه جزء تشکیلشدهاند:
یونهای +٣ Fe و+2 Fe
یک کمپلکس همچون EDDHA،DTPA،EDTA، اسیدهای آمینه، اسیدهای هیومیک – فولویک و سیترات
سدیم (+Na) و یا آمونیوم (+٤NH)
کلاتهای مختلف میتوانند یونهای آهن را در pH مای مختلف باقدرتهای متفاوتی نگهدارند. همچنین حساسیت آنها به جایگزینی یونهای رقیب با یون آهن نیز متفاوت است. بهعنوانمثال، هنگامیکه غلظت یونهای کلسیم یا منیزیم زیاد باشد ممکن است این یونها بهجای آهن کلات شده بنشینند. البته واکنش اهن +2 Fe با عامل کلات کننده تحت شرایط سختی انجام می شود بنا براین این محصولات در بازار به سختی یافت میشوند و اغلب به رنگ سبز لجنی می باشند.
Fe-EDTA
این کلات آهن در pH کمتر از ۶ باثبات بوده اما در pH بالاتر از ٥/٦ حدود ٥٠% آهن آن برای گیاه غیرقابلدسترسی میشود؛ بنابراین مشخص است که این کلات در خاکهای قلیایی غیرقابلاستفاده است. همچنین این کلات میل ترکیبی زیادی با کلسیم دارد و به همین دلیل توصیه میشود تا از مصرف آن در خاکها و یا آبهایی که مقدار کلسیم آنها بالا است خودداری گردد. لازم به ذکر است که EDTA یک کلات بسیار پایدار برای عناصر غذایی کممصرف، بهجز آهن است که میتواند pHهای بسیار بالا را نیز تحمل کند.
Fe-DTPA
این کلات ثبات خود را تا pH حدود ۷ حفظ کرده و حساسیت آن به جایگزین شدن کلسیم بهجای آهن کمتر است.
Fe-EDDHA
این کلات آهن حتی تا pHهای بیشتر از ۱۱ نیز باثبات بوده و به همین دلیل نیز گرانترین نوع کلات آهن موجود در بازار است. کلاتهای EDDHA ممکن است به دو صورت اورتو – اورتو و یا اورتو – پارا باشند. هرچه میزان اورتو -اورتو در یک کلات آهن EDDHA بیشتر باشد، آن کود آهن مرغوبتر بوده، پایداری کلات آن بیشتر است و به طبع گرانتر نیز است. شکلهای زیر آرایش فضایی اورتو – اورتو و اورتو – پارا را نشان میدهند.
امینو کلات اهن
این کلات آهن تا pHهای بیشتر از 9 نیز باثبات بوده و به دلیل ساختار طبیعی و شناخته شده توسط گیاه و سایز کوچکتر بهترین نوع این کلاتها می باشد . کلاتهای بر پایه اسید امینه ممکن است به دو فرم اهن دو و یا سه ظرفیتی باشد.و همچنین میتواند با یک و یا چندین نوع اسید امینه کلات شده باشد. هرچه تعداد انواع اسید امینه در ساختار امینو کلات زیاد تر باشد این محصول دارای تعداد بیشتری از انواع کلات های امینو اسید است بنابر این مرغوبیت و کارایی بالاتری از خود نشان میدهد. هرچه تعداد انواع L اسید امینه یک کلات امینو کلات آهن بیشتر باشد، آن کود آهن مرغوبتر بوده، پایداری کلات آن بیشتر است و به طبع گرانتر نیز است.
نتیجه گیری
به دلیل سایز مولکولی کوچکتر (در حدود نانو ) فرم طبیعی و هماهنگ با ساختار سلولی گیاهی ، استفاده از انواع اسید امینه در ساختار که هم موجب افزایش جذب آهن و هم مزایای اسید امینه را دارد و عدم تجمع ماده مضر EDTA داخل سلول و …. در مقام مقایسه انواع کلات های اهن میتوان گفت :بهترین و مرغوبترین کلات آهن از نوع امینو کلات می باشد.
بهداد، ابراهیم. ۱۳۵۸. بیماریهای درختان میوه در ایران، چاپ نشاط اصفهان.
فیلسوف، فریدون. ۱۳۶۵. بررسی اثر کودهای آهندار بر معالجه زردی ناشی از کمبود آهن برگ درختان به در اصفهان. ششمین کنگره گیاهپزشکی ایران دانشگاه صنعتی اصفهان.
صفری، محمد. ۱۳۹۰. مبانی بیوشیمی کشاورزی. انتشارات دانشگاه تهران.
فیزیولوژی و چگونگی جذب عناصر غذایی از طریق محلول پاشی
جذب عناصر غذایی در گیاهان اغلب از دو مسیر برگی و ریشه ای انجام میگردد در این مقاله سعی داریم به صورت مختصر فیزیولوژی جذب برگی در گیاها را شرح دهیم در ادامه با ما باشد.
تغذیه برگی (محلولپاشی)
کلیه اندامهای گیاهان اعم از ریشه، ساقه، شاخه، برگ و حتی میوه میتوانند آب، گازها و مواد غذایی را جذب یا دفع کنند. اطلاعات موجود درباره جذب مواد غذایی از راه برگ قسمتهای قابلتوجهی از مطالعات تغذیه گیاهی را تشکیل میدهد. تأمین مواد غذایی از راه برگ و شاخه، تغذیه برگی، کودپاشی برگی یا کودپاشی هوایی نامیده میشود.
مزایا و معایب تغذیه برگی (محلولپاشی)
بهطورکلی استفاده از هر روشی و به هر منظوری ممکن است مزایا و معایبی داشته باشد که موفقیت در کاربرد آن بستگی به فراهمآوردن زمینه لازم و درنظرگرفتن تمامی فاکتورهای مؤثر و برآیند مزایا و معایب استفاده از آن روش را دارد.
مزایا
با محلولپاشی ترکیبهای غذایی واکنشهای تشکیل کمپلکس در خاک، مخصوصاً در مورد عناصر کممصرف حذف میشود.
از میزان مصرف کود و ترکیبهای غذایی در واحد سطح کاسته میشود که علاوه بر صرفهجویی در هزینهها از آلودگی و شوری خاک ناشی از مصرف زیاد کودها، بهویژه در مناطقی که آب یا خاک شور باشد، جلوگیری میکند.
معمولاً گیاه درنتیجه حمله آفات یا بیماریها قسمتی از نیروی رشد و نمو خود را از دست میدهد. مصرف توأم سم و کود هزینههای محلولپاشی را کاهش داده و علاوه بر اینکه عامل زیانبخش را برطرف میکند مواد غذایی را نیز سریع به گیاه میرساند، بنابراین گیاه تقویت میشود.
پاسخ گیاه در مقابل محلولپاشی عناصر غذایی خیلی سریع است و میتوان در زمانهای مختلف از آن استفاده نمود.
در طول مرحله زایشی در اثر رقابت برای جذب کربوهیدراتها بین اندامهای زایشی (دانه و میوه) و ریشهها، از فعالیت ریشه کاسته میشود درنتیجه جذب مواد غذایی کاهش مییابد. در این مرحله محلولپاشی عناصر غذایی این رقابت را کاهش میدهد. بهخصوص در محصولاتی مانند درختان میوه، گندم، جو و بقولات در مرحله زایشی بین رشد و نمو میوهها و رشد ریشهها رقابت شدیدی به وجود میآید.
در باغات کشور در اثر عوارضی ناشی از کمبود کلسیم در میوه، مثل لکه تلخ، لهیدگی، پوسیدگی گلگاه و ترک برداشتن میوه بهوفور یافت میشود. ولی باوجود وفور این عنصر در خاک، جذب آن به دلایل متعددی مثل کند بودن حرکت آن در داخل گیاه و انتقال بهطرف میوهها با اشکال روبهرو میگردد، لذا بهمنظور تأمین کلسیم و منیزیم در درختان میوه بهترین را محلولپاشی است.
در خاکهای آهکی به دلیل بالا بودن pH، آهک و مصرف کودهای فسفره بیش از نیاز، جذب آهن و منگنز معمولاً دچار اشکال میگردد و در بیشتر موارد کمبود آهن و روی دیده میشود؛ بنابراین محلولپاشی در چنین شرایطی مؤثرتر و باصرفهتر از مصرف کلاتهای آهن و روی گرانقیمت در خاک است.
معایب
در صورت بکار بردن غلظتهای بالا و عدم توجه به خصوصیات گیاه و ترکیب غذایی مورداستفاده، ممکن است آسیبدیدگی برگ و میوه به وجود آید که در این صورت خسارتهای جبرانناپذیری به درخت و محصول وارد میشود.
راههای جذب برگی
1.جذب کوتیکولی
یکی از راههای جذب عناصر، جذب آنها از راه کوتیکول است. وقتیکه مواد غذایی روی سطح برگ قرار میگیرند. تا وارد سلولهای گیاهی شوند مدتزمانی طول میکشد. این زمان را زمان کمون میگویند. هرچه این دوره کوتاهتر باشد پاسخ گیاه به عناصر غذایی محلولپاشی شده سریعتر خواهد بود. مدتزمان کمون بستگی به ضخامت کوتیکول دارد.
2.جذب روزنهای
جذب از راه روزنهها اهمیت بسزایی دارد. چون سلولهای محافظ روزنه قدرت جذب بالایی دارند و اگر محلول به درون روزنه نفوذ کند، مقدار زیادی از آن جذب خواهد شد. جذب از داخل حفره زیر روزنه وقتی اهمیت پیدا میکند که به محلول مورداستفاده مویان یا ماده خیسکننده اضافه شود تا نفوذ محلول به داخل روزنه امکانپذیر باشد .
ازآنجائیکه ساعات خنک روز منطبق بازمان بازبودن روزنههای برگ است انجام محلولپاشی در این ساعات باعث افزایش راندمان جذب برگی از طریق روزنهها میشود .
3.جذب از طریق کرکهای کوتیکولی
کرکهای کوتیکولی نیز میتوانند به نفوذ بخشی از مواد غذایی محلولپاشی شده کمک کنند. اهمیت این مسیر در جذب برگی به گسترش و موقعیت این کرکها بستگی دارد که خود از مشخصههای بلوغ برگ و گونه گیاه به شمار میرود .
چگونگی جذب و انتقال ترکیبهای محلولپاشی شده
روند جذب برگی تفاوت زیادی با عمل جذب توسط ریشه ندارد. بدین معنی که در اینجا نیز پدیده جذب به دو صورت فعال و غیرفعال وجود دارد .
گرفتن عنصر بهوسیله گیاه مرحله اول جذب است. مرحله دوم، نفوذ عنصر، یا یون به داخل سلولهای سطحی است. برعکس مرحله اول که کمتر تحتتأثیر عوامل خارجی قرار میگیرد این قسمت از جذب که درواقع جذب غیرفعال است تحتتأثیر رطوبت نسبی محیط و درجه حرارت قرار میگیرد. مرحله سوم جذب از راه برگ، این قسمت از جذب نیز تابع تأثیر عواملی ازجمله عواملی کندکننده، میزان اکسیژن و pH است. درنتیجه با شستشوی برگها عناصر جذبشده شسته نمیشوند. وابستگی جذب برگی به عوامل ذکرشده نشان میدهد که حداقل یک مرحله از جذب برگی یونها تابع جذب فعال است.
بنابراین هرچقدر درخت در شرایط رشدی مطلوبتری قرار گرفته باشد و فعالیتهای حیاتی آن بهتر صورت گیرد راندمان جذب فعال عناصر محلولپاشی شده بیشتر خواهد بود .
مسیر انتقال عناصر بعد از نفوذ آنها به کوتیکول
1.از طریق فضاهای آزاد بین سلولی
بعضی از مواد و یونها ممکن است از طریق این فضاهای آزاد بین سلولی که مسیر اپوپلاسمی نامیده میشود وارد سلولهای پارانشیمی شده و پس از تبدیلشدن به مواد غذایی در طی عمل فتوسنتز از طریق آوند آبکش به سایر قسمتهای درخت منتقل گردند .
2.از طریق غشاءهای سلولی
بعضی از یونها ممکن است پس از جذب سطحی بهوسیله فرایند جذب فعال از غشاء سلولهای اپیدرمی و پارانشیمی عبور کرده و ازآنجا به سیتوپلاسم سلولها و سپس سایر قسمتهای گیاه منتقل شوند .
3.حرکت یونها از راه پلاسمودسماتا
پلاسمو دسماتا منافذ ریزی هستند که ارتباط بین سلولهای گیاهی همجوار را برقرار میسازند. برخی از یونها بهوسیله این منافذ از سلولهای سطحی به سلولهای پارانشیمی و درنهایت به سیستم آوند انتقال مییابند .
عوامل مؤثر بر جذب برگی عناصر غذایی
عوامل مؤثر بر جذب عناصر غذایی از طریق برگ عبارتاند از: نور، درجه حرارت، رطوبت نسبی، سن برگ، حالتهای تغذیهای برگ، فرم شیمیایی عنصر مورداستفاده و مواد خیسکننده
1.نور
بسته به گونه گیاه ممکن است نور اثرات متفاوتی داشته باشد. میزان کوتین و یا مومهای کوتیکولی و ضخامت کوتیکول، در شدت نور زیاد بیشتر از شدتهای پایین، در گیاهان کلم، اکالیپتوس، غلات و میخک است.
2.درجه حرارت هوا
درجه حرارت هوا نیز بر جذب برگی عناصر غذایی از سطح برگ، اثرات متفاوتی دارد اثر دما روی توسعه مومها بستگی به گونه گیاه دارد.
3.رطوبت نسبی
این مسئله ازنظر کاربردی حائز اهمیت است که محلولپاشی در موقعی از روز انجام شود که رطوبت نسبی بالا باشد. بدین منظور توصیه میگردد که محلولپاشی در صبح یا عصر انجام شود. انجام آبیاری قبل یا بعد از عمل محلولپاشی باعث افزایش رطوبت محیط و درنتیجه افزایش جذب برگی میشود .
4.سن برگ
بهطورکلی بین سن برگ و میزان جذب برگی، ترکیبهای غذایی، ارتباط وجود دارد .
5.حالتهای تغذیهای گیاهی
زمانی که گیاه دچار کمبود عنصری باشد با محلولپاشی آن عنصر جذب بیشتر و سریعتر انجام میشود. هرچقدر فعالیت متابولیکی گیاه بیشتر باشد جذب فعال عناصر که مستلزم صرف انرژی میباشند از طریق برگ نیز بیشتر میگردد؛ بنابراین یکی از اثرات وضع مناسب تغذیهای گیاه در بالابردن جذب برگی عناصر از این طریق اعمال میشود .
6.فرم شیمیایی عنصر مورداستفاده
از مهمترین عوامل مؤثر در گزینش موادی که برای محلولپاشی مصرف میشوند خطر سوختگی برگ و میزان جذب عنصر یا عناصر موجود در ترکیب است. اثر هر محلول بر سوختگی برگی با تفاوت فشار اسمزی بین محلول مصرفشده و شیره سلولی مشخص میگردد. اگر فشار اسمزی محلول بیش از فشار اسمزی شیره سلولی باشد، آب از بافت گیاهی خارج گشته و سوختگی حاصل میشود .
فرمهای کلاته شده عناصر غذایی مخصوصاً کلاتهای آهن، کارایی آنها در مقایسه بانمکهای معدنی (احتمالاً بهخاطر تحرک زیادشان در گیاه) بسیار بالاتر است .
7.pH محلول
pH محلول در جذب عناصر غذایی معین توسط برگ و میوهها مؤثر است بهطورکلی در گیاهان مختلف جذب عناصر غذایی در pHهای متفاوت صورت میگیرد
اثرات محلولپاشی عناصر غذایی روی خصوصیات محصولات باغی
1.رشد رویشی
بهطورکلی اگر محدودیت رشد رویشی درختان درنتیجه کمبود عناصر غذایی به وجود آمده باشد، با محلولپاشی این عناصر رشد رویشی تحریک میشود. ولی درصورتیکه وضعیت عناصر غذایی در درخت مناسب باشد پاسخ رشدی گیاه نسبت به محلولپاشی عناصر موردنظر کم خواهد بود. ازجمله عناصری که در کنترل رشد رویشی درخت نقش مهمی دارد ازت است. در مورد محلولپاشی کود ازته در ارتباط با رشد رویشی درختان میوه بسته به وضعیت ازت درخت چند نکته باید در نظر گرفته شود .
2.زمان محلولپاشی
در درختان بارور پس از آغاز رشد در بهار یک مرحله رشد فعال رویشی وجود دارد و سپس این رشد متوقف میشود. از طرف دیگر در اکثر درختان مناطق معتدل معمولاً تشکیل جوانههای گل در سال قبل از بازشدن شکوفهها اتفاق میافتد و معمولاً گل انگیزی پس از توقف رشد رویشی صورت میگیرد. برای مثال در سیب گل انگیزی جوانهها در خردادماه صورت میگیرد، در این زمان رشد رویشی نامتعارف باعث میشود که مواد غذایی صرف تولید شاخوبرگ اضافی شود و این شاخوبرگ اضافی بر سر عناصر غذایی، با جوانههای در حال تمایز رقابت میکنند؛ بنابراین محلولپاشی اوره یا سایر ترکیبات ازته باید زمانی صورت گیرد که باعث تحریک رشد رویشی نگردد و درصورتیکه ذخیره ازته درخت خیلی کم نباشد ترجیحاً بعد از توقف رشد رویشی شاخهها محلولپاشی انجام گیرد .
3.منبع کودی
در انتخاب منبع کودی برای تأمین عناصر غذایی بهغیراز ازت در درختان بارور درصورتیکه ذخیره ازته مناسب باشد و محلولپاشی در اول فصل ضرورت داشته باشد باید از منبعی استفاده شود که یون همراه عنصر موردنظر ازت نباشد .
4.سن درخت
اگر درخت در سنین اولیه رشد و در دوره نونهالی قرار داشته باشد استفاده از منبع ازته سبب تحریک رشد رویشی میگردد و گیاه این مرحله را سریعتر پشت سر گذاشته و زودتر به مرحله بلوغ و باروری خواهد رسید. ولی در درختان بارور مخصوصاً در ارقامی که قدرت رشد رویشی آنها زیاد است در استفاده از ازت باید جانب احتیاط را رعایت نمود .
5.میزان محصول
بهطورکلی تأثیر عناصر غذایی بر میزان محصول یک درخت از طریق اثرات آنها روی تشکیل میوه، اندازه میوه و جلوگیری از ریزش میوه اعمال میشود.
6.کیفیت محصول
کلسیم ازجمله عناصری است که در ترکیب شیمیایی و ساختمانی دیواره سلولهای گیاهی نقش بسیار مهمی را دارد؛ بنابراین بالابود میزان کلسیم دریافت میوه میتواند بهاندازه زیادی از اتلاف میوهها در زمان برداشت در طی نگهداری آنها در سردخانه جلوگیری نماید. همچنین علاوه بر پایین بودن سطح کلسیم در میوه بالا بودن نسبتهای پتاسیم به کلسیم (K/Ca) و میزیم به کلسیم (Mg/Ca) نیز در ظهور بعضی از بیماریهای فیزیولوژیک مؤثر است. کمبود بُر سبب توسعه عارضه چوبپنبهای شدن میوه میشود. بعضی مواقع بین علائم کمبود بُر و چوبپنبهای شدن و لکه تلخ میوه شبهههایی به وجود میآید زیرا چنین علائمی باهم مشابه هستند. بُر علاوه براثر مستقیم روی کیفیت میوه، میتواند تحرک کلسیم را نیز در درخت افزایش دهد به طور غیرمستقیم سبب بالارفتن کیفیت میوه سیب گردد
سرعت جذب عناصر غذایی
جدول زیر سرعت نسبی جذب عناصر غذایی برحسب مدتزمان لازم برای جذب 50% از یونهای برگ پاشی شده مورد مقایسه قرار گرفته است.
چندین دهه است که کشاورزان از آمینو اسیدها بهصورت محلولپاشی و مصرف خاکی استفاده میکنند. کشاورزان در کشورهای پیشرفته که با کمبود مواد معدنی ارگانیک و گاها ساختار ضعیف مواد خام و با استفاده بیشازحد از اکوسیستم موجب ضعیف شدن خاک شده اند، به مزایای آنها بهعنوان تقویتکنندههای ارگانیک و محرکهای زيستی پی بردهاند.
آمینواسیدهای ضروری، استاندارد و غیرضروری
تمام پروتئینها تركيبات مولكولی با صدها آمینو اسید مختلف بهعنوان بلوکهای سازنده بنيادين میباشند. حدود 500 آمینو اسید شناساییشدهاند اما اغلب دانشمندان اتفاقنظر دارند كه ٢٠ (برخی میگویند ٢١ يا ٢٣) اسید آمینه استاندارد موردنیاز برای حيات وجود دارد.
در بيوشيمی پروتئين، واژه آمینو اسیدهای ضروری كاملاً رايج است. اين در اشاره به نيازهای بشر است زيرا بدن انسان قادر به توليد آنها نيست و بايد آنها را از رژيم غذايی خود به دست آورد. در مقابل، اغلب گياهان قادر به تركيب و تولید آنچه نياز دارند هستند. بااینحال، اين فرایند نياز به انرژی فراوان دارد بنابراين ايده خوبی است كه آن را از آمینو اسیدهای آماده استفاده از طريق تغذيه برگی يا ريشه فراهم كرد.
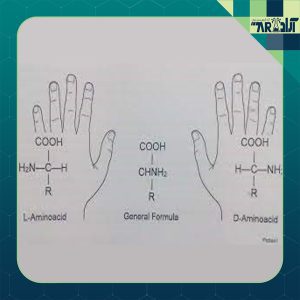
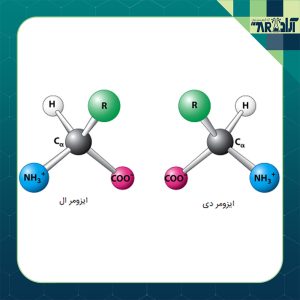
آمینواسیدهای چپگرد در مقابل راستگرد
يكی از مهمترین مفاهيم آمينواسيدها درك تفاوت بين L آمینواسیدها چپگرد و D آمینو اسیدها راستگرد است. اين موضوع تا حدودی تخصصی بوده ولی تا حد امکان این موضوع را برای شما در این مقاله بیان خواهیم نمود.
آنچه ما از آن اطلاع داريم اين است كه طبيعت (حيات) تصميم گرفته است تنها از يك نوع آمینو اسید استفاده كند. بهجز موارد استثناء بسيار نادر، طبيعت تنها از نوع چپگرد يا L آمینو اسید استفاده میکند. اين اثر بهعنوان كايرالی (تصوير آيينه) شناختهشده و به تمام آمینو اسیدهای فعال اپتيكی حاوی يك آلفا گروه آمينو برای اسید کربوکسیلیک اطلاق میشود.
پيكربندی طبيعی در سری آمینواسیدها، L ناميده میشود (آمینو اسیدهای چپگرد). در اينجا “L مخفف”Laevo” (به معنی سمت چپ) است، نه “Left (به معنی چپ). نوع دیگر D آمینو اسیدها هستند كه در آن “D مخفف”Dextro (به معنی سمت راست) است. هر دو آنها دارای ساختار مولكولی دقيقاً مشابه بوده و تصاوير آیینهای دارند. بهترين راه برای فهميدن تفاوت، دستان خودتان است. ممكن است دستها مشابه باشند – استخوانها، رگها، انگشتان – اما نمیتوانید دستكش دست چپ را در دست راست بپوشيد.
برای استفاده آمینو اسیدها در حيات، اين مولکولها بايد مناسب دریافتکنندهها باشند. به اين دليل است كه يك ارگانیسم دست چپ نمیتواند از آمینو اسید های دست راست استفاده كند. بنابراین، L آمینو اسید ها تنها از فرایندهای ارگانيك طبيعی، ازجمله آبكافت آنزيمی (شكستن پروتئینها بهوسیله آنزيم) یا قلیایی و یا اسیدی گرفته میشوند. در سنتز های مصنوعی تولیدکننده D و L آمینو اسید میباشند كه ممكن است در كاربردهای ديگر سودمند باشند، ولی برای حيات خیر.
آبكافت آنزيمی فرایندی است كه در آن آنزیمها و مواد ارگانيك با آب و گرما در كنار يكديگر قرار میگیرند. مواد ارگانيك، با آبكافت تجزیهشده و به عناصر بنيادين تبديل میشوند (L آمینو اسیدها)؛ مانند هضم کربوهیدراتها، چربی و پروتئینها در انسان. اين فرایند هزينه بالايی داشته و نيازمند تجهيزات خاصی است اما تضمینکننده L آمینواسیدهای دوستدار حيات و خالص است.
L آمینو اسیدها و کاربرد آنها
در محيط خاكی طبيعی با اكوسيستم سالم و شكوفا، گياهان آمینواسیدها را از تجزيه مواد ارگانيك به دست میآورند. گياه در زمان نياز آنچه لازم دارد را توليد میکند. متأسفانه، كشاورزی مدرن حيات خاك سالم را تضعيف و نابود كرده و سطح تشكيل طبيعی آمینو اسیدها را كاهش داده است. بنابراين، گياهان نياز به افزايش انرژی در توليد آمینو اسید دارند كه از صرف انرژی در كاربردهای سودبخشتر (افزایش کیفیت و کمیت محصول )جلوگيری میکند.
پيش از اينكه آمینو اسیدها بتوانند كاربرد ضروری در گياهان داشته باشند، بايد ازنظر زيستی در دسترس باشند. تنها اعمال آنها در گياه كافی نيست. آمینو اسیدها برای جذب از طريق ريشه يا بافت بايد يا عاری از پپتيدهای مولكولی (دی پپتيد، تری پپتيد) بوده يا تعداد كمی از آن را داشته باشند. آمینو اسیدهایی كه در زنجیرههای بلند تركيب میشوند مولکولهای بسيار بزرگ بوده و قادر به نفوذ در بافت گياه نيستند. بهصورت تئوری، مولکولهای كوچك دارای وزن مولكولی كمتر از ٩٠٠ دالتون هستند كه اجازه پراكندگی سريع در تمام غشاها و رسيدن به محلهای درونسلولی رادارند.
كاربردهای آمینو اسیدها در متابوليسم گياهان تقريباً نامتناهی است و علم به طور متناوب در حال كشف متابوليسم بيشتر است.
عملکرد آمینواسیدها در گیاه
افزايش توليد كلروفیل
ارائه منبع غنی از نيتروژن ارگانيك.
تحريك توليد ویتامینها
تأثیر بر سیستمهای آنزيمی مختلف
تحريك گلدهی
حمايت از محيط ميوه
افزايش محتوی مغذی، اندازه، طعم و رنگ میوهها
افزايش كيفيت محصول
افزايش مقاومت در برابر پاتوژنها و آفتها
حمايت از گياه در برابر فشارها و استرسهای زنده (آفات و ویروسها) و غیرزنده (سرمازدگی, کمآبی, سموم و غيره).
سنتز پروتئین
آمینو اسیدها ساختار بنيادين پروتئینها میباشند. آمینواسیدهای استاندارد بهصورت نامتناهی تركيب میشوند تا پروتئینهای بیشماری توليد نمايند. اين پروتئینها برای بسياری از عناصر ساختاری بافت گياه ضروری هستند.
پروتئینها كاربردهای فراوانی دارند: ساختاری (حمايتی)، متابوليك (آنزیمها و تحريك)، انتقال ریزمغذیها، حفظ آمینو اسید. درواقع، پروتئینها تقريباً با تمام فرایندهای زيستی سازگارند. گياهان، برخلاف انسانها، پروتئینهای خود را بر اساس مرحله خاص رشد، نيازهای تغذيه، فشار و غيره توليد مینمایند. بااینوجود، دو موضوع اهميت فوقالعادهای دارند:
گياهان تنها در صورتی قادر به ايجاد پروتئینهای موردنیاز هستند كه مواد خام وجود داشته باشد.
توليد آمینواسیدها يك فرایند بسيار انرژی بر است.
بنابراين، تولید L آمینو اسیدهای اضافی در ریشه ها يا بافت برگ ها، گياه مواد فراوانی ذخیره کرده و برای ايجاد پروتئینهای مهم انرژی ذخيره میکند.
نقش آمینو اسیدها در مقاومت گیاهان برابر استرسهای غیرزیستی
استرس غیرزیستی، ازجمله دمای بالا/پايين، خشكی، حملات آفتها، بيماری، يا فيتوتاكسيك در اثر استفاده از آفتکشهای شيميايی آثار منفی بر متابوليسم گياه دارند. به طور حتم اين امر موجب كاهش كيفيت و ميزان محصول میشود. استفاده از کود آمینو اسید قبل یا زمان و پس از شرايط استرسزا به گياهان موادی میدهد كه به طور مستقيم پیشگیریکننده بوده و اثر بازسازی دارد.
زمانی كه گياه تحت استرس است، خود توليدی آمینو اسیدها كاهش مییابد زيرا این یک پروسه انرژی بر است. در عوض، گياه پروتئینهای موجود را آبكافت (شكستن) میکند تا به آمینو اسیدهای موردنیاز دست يابد. اين فرایند انرژی كمتری نسبت به توليد آمينو اسيدها نياز دارد. همچنين اين بدين معنی است كه گياه ممكن است خود را “خود خواری” كند، مگر اينكه آمینو اسید هایی از طريق مكمل فراهم گردند.
نقش آمینواسیدها در فتوسنتز گیاه
فتوسنتز مهمترین فرایند بيوشيمی گياه است. يك گياه به توليد قند از دیاکسیدکربن، آب، و انرژی نور میپردازد. پس از آن قندهای جمعآوریشده (کربوهیدراتها) توسط گياه بهعنوان منبع انرژی فرایندهای متابوليك ديگر استفاده میشوند. آمینواسیدها تأثیر فراوانی در اين عملكرد متابوليكی مهم دارند.
L- گليسين و L- گلوتاميك متابولیتهای ضروری برای توليد كلروفيل و تشكيل بافت هستند. اين آمینو اسیدها موجب افزايش غلظت كلروفيل در گياهان میشوند. كلروفيل بيشتر, به معنی افزایش جذب انرژی است كه موجب افزايش فتوسنتز خواهد شد.
نقش آمینو اسیدها در گردهافشانی و تولید میوه
گردهافشانی و توليد ميوه دو تا از مهمترین مراحل گياهان هستند. آمینو اسیدها در زمان اوج فعالیتهای متابوليكی حائز اهميت هستند:
L- هيستيدین به رسيدن ميوه كمك میکند.
L- پرولين موجب افزايش فعاليت گردهافشانی میشود
L- ليسين، L- متيونين و L- گلوتاميك اسيد موجب افزايش توليد گرده و طول لوله آن میشود.
L- الانين، L- والين و L- لوسين موجب افزايش كيفيت ميوه میشوند.
نقش آمینو اسیدها در فعالیت باکتری ها و رشد گیاه
ازآنجا که تمام حيات بستگی به آمینو اسیدها دارد، اين شامل تمام باکتری های موجود در حول ناحيه ریشه میشود. باکتری ها تا جایی که میتوانند از آنها استفاده میکنند. برخی از آنها بهعنوان بلوکهای سازنده برای عناصر سازنده و توليد پروتئين شركت میکنند. برخی ديگر بهعنوان محرك توليد عناصر رشد و هورمون مختلف استفاده میشوند. برای مثال، L- متيونين يك ماده تشکیلدهنده عوامل رشد است كه غشاهای سلول در باکتری را تثبيت میکند. همچنين برخی از باکتری ها آمینوها را بهعنوان منبع پروتئين و نيتروژن مصرف مینمایند. بهعلاوه، آمینو اسیدها در خاك يك منبع غنی مواد ارگانيك برای كمك به ايجاد سازه، حاصلخيزی و حفظ آن را فراهم میکنند.
آمینو اسیدها بهعنوان منبع ثابت نیتروژن ارگانیك
رایجترین اشكال شناختهشده نيتروژن مورداستفاده توسط گياهان نیتراتها –(NO٣) و آمونيوم +(NH٤)هستند. فراهمکردن نيتروژن بهعنوان تغذيه دشوار است، زيرا يك گاز طبيعی است و بهآسانی از خاك آزاد میشود. بيشتر كودهای تجاری حاوی اين دو شكل باکیفیت بالا هستند. گياهان از هر دو شكل استفاده نمینمایند، هرچند گياهان مختلف موارد ترجيحی متفاوتی دارند. بااینوجود، يك منبع ديگر نيتروژن وجود دارد. اين منبع كمتر موردبحث قرار میگیرد اما همچنان يكی از فاكتورهای مدنظر در كشاورزی بيولوژيكی و ارگانيك در اروپا است. مواد ارگانيك مانند آمینو اسیدها حاوی نيتروژن هستند. زمانی كه گياه آمینواسید خود را تخليه میکند از نيتروژن ارگانيك استفاده مینماید.
ازآنجاکه بخشی از نيتروژن در گياه برای توليد پروتئين و آمینو اسید استفاده میشود، با فراهم نمودن شكل آماده، گياه نياز كمتری به نيترات و آمونيوم برای اين فعالیتها دارد.
چرا اين موضوع حائز اهميت است؟ مانند هر چيز ديگری، استفاده بیش از حد از يك جنبه موجب بروز مشكلاتی در جای ديگر میشود. نیتراتهای فراوان به طور ويژه تمايل به ايجاد رشد سريع و كشيدگی سلول دارند؛ مانند شكل سريع سلولهای درحالرشد، ديواره سلول كشيده و نازك میشود. اين بافت نازک يك هدف عالی برای آفتهای حملهکننده است. اين پديده را میتوان با محصولات زمينی زيادی مانند ذرت مشاهده كرد . نيترات فراوان نيز موجب ناسازگاری در برابر ديگر مواد معدنی مهم مانند كلسيم، منیزيم و پتاسيم میشود.
زمانی كه نیتراتها در حالت متعادل باشند، فراهم نمودن نيتروژن ارگانيك میتواند موجب رشد سلولها با شكل طبيعی و محكم شود. اين امر موجب ايجاد يك گياه قویتر با سلولهای سالم با مقاومت بيشتر در برابر استرس و حملات میشود.
در زمان صحبتکردن از سلولهای گياهی و آمینو اسیدها، نمیخواهیم حمايت از آنها را برای تنظيم تعادل تراوشی فراموش كنيم. روزنههای هوايی ساختارهای سلولی هستند كه تعادل آب گياه را كنترل مینمایند. همچنين آنها در طول تعرق (“تنفس” از برگها) و همچنين جذب ریزودرشت مغذیها استفاده میشوند. بازگشتهای روزنه هوايی با عوامل خارجی (نور، رطوبت، دما، و غلظت نمك) و داخلی (آمینواسیدها، پتاسيم در دسترس و غيره) كنترل میشوند. روزنههای هوايی در زمان نور و رطوبت كم بسته میشوند؛ همچنين اين اتفاق در زمان دما و غلظت نمك زياد نيز رخ میدهد. بستهشدن آن موجب كاهش فتوسنتز و تعريق و افزايش تنفس میشود. اين امر موجب كاهش تعادل متابوليك و کمشدن سرعت رشد میشود. گلوتاميك اسيد بهعنوان يك عامل تراوشی برای سلولهای حفاظتی عمل میکند كه میتواند بازشدن روزنه را افزايش دهد.
شما می توانید تاثیر اسید آمینه را بر روی گیاه گندم را در صفحه اینستاگرام شرکت مشاهده کنید.
آمینو اسیدها بهعنوان عامل کلاته کننده
يكی از قابلتوجهترین نقشهای آمینو اسید ها افزايش تنوع زيستی مغذیها است. گياهان به دليل ساختار مولكولی و شارژ يونی، برخی از مغذیها را جذب نمیکنند. آمينو اسيدها (و برخی ديگر از اسيدهای ارگانيك) اين مواد معدنی غیرقابلدسترس را قابلجذب و انتقال در بافتهای گياه میکنند.
با كلات با آمینو اسیدها، مقدار كلی مواد معدنی محلول قابل جذب موجود در محلول مغذی افزایش یافته و به واسطه امینو اسید جذب و انتقال از طریق گیاه افزایش می یابد. بهعلاوه، آمینواسیدها اجازه تغذيه برگی سودبخش با انتقال مواد معدنی توسط روزنههای هوايی را میدهند.
اسيدهای L- گليسين، L- گلوتاميك و L- آسپارتيك بهعنوان عوامل كلات كننده بسيار سودمند شناخته میشوند كه دليل عمده آن وزن مولكولی كوچك آنها است. وزن آنها اجازه حركت سريع در غشاهای سلول را میدهد. علاوه بر افزایش دسترسی مواد مغذی خوب، آمینواسیدها برای كاهش سميت فلز در گياهان و خاك با پيوند با فلزهای سنگين شناخته میشوند. اين امر به ايجاد تعادل سطوح عناصر گوناگون در واسطه كمك میکند.